First of all, no. I am not the lovechild of star-crossed siblings, or even cousins, or even second cousins.
This is a gee-whiz kind of post. But the issues are not insignificant.
Hear me out with the background, first, before I get to the part where my eyes bug out of my head and I pull out my kid's Crayola box and start drawing.
If you've learned about sociobiology, or evolutionary psychology, or inclusive fitness, or kin selection, or the evolution of cooperation and even "altruism," or if you've read The Selfish Gene, or if you've been able to follow the debate about levels of selection (which you can peek at here)...
... then you've heard that you're related to your parents by 1/2, to your siblings by 1/2 as well, to your grandparents and grandchildren by 1/4, to your aunts and uncles and nieces and nephews by 1/4 as well, and to your first cousins by 1/8 and so on and so forth. (Here's some more information.)
So, for example. For evolution (read: adaptationism) to explain how cooperative social behavior could be adaptive in the genetic sense, we use the following logic provided by Bill Hamilton, which became known as "Hamilton's Rule":
The cost to your cooperation or your prosocial behavior (C) must be less than its benefit to you (B), reproductively speaking, relative to how genetically related (r) you are to the individual with whom you're cooperating. That could have come out smoother. Oh, here you go:
C < rB, or B > C/r
If you're helping out your identical genetic twin (r=1.0), then as long as the benefit to you is greater than the cost, it's adaptive.
C < B, or B > C
If you're helping out your daughter (r = 0.5) then as long as the benefit to you is greater than twice the cost, it's adaptive.
C < (1/2)B, or B > 2C
So already, the adaptive risk to helping out your daughter or your brother is quite higher. And it's even harder to justify the cooperation between individuals and their sibs' kids, and grandkids, especially ESPECIALLY non-kin. But, of course creatures do it! And so do we.
As relatedness gets more distant and distant, we go from 2 times the cost, to 4 times, 8 times, 16, 32, 64 etc... You can see why people like to say "the math falls away" or "drops off" at first or second cousins when they're explaining where the arbitrary line of genetic "kin" is drawn. If you offer up a curious, "we're all related, we're all kin," someone out of this school of thought that's focused on explaining the evolution of and genes for social behavior may clue you in by circumscribing "kin" as the members of a group that are r = 1/8 or r = 1/16 but usually not less related than that.
This has long bothered me because we're all genetically related and so much cooperation beyond close kin is happening. And it's been hard for me, as someone who sees everything as connected, to read text after text supporting "kin selection" and "kin recognition" (knowing who to be kind to and who to avoid bleeping), to get past the fact that we're arbitrarily deciding what is "kin" and it seems to be for convenience. I'm not doubting that cooperation is important for evolutionary reasons. Quite the contrary! It's just that why is there so much math, based in so many potentially unnecessary assumptions about genes for behavior, gracing so many pages of scientific literature for explaining it or underscoring its importance?
(It could just be that as an outsider and a non-expert I just don't understand enough of it and if I only did, I wouldn't be gracing this blog with my questions. But let's get back to my reason for posting anyway because it's potentially useful.)
Right. So. Even for folks who aren't part of evolution's academic endeavor, it's obvious to most that we're one half dad and one half mom. The sperm carries one half of a genome, the egg another, and together they make a whole genome which becomes the kid. Voila!
There's even an adorable "Biologist's Mother's Day" song about how we've got half our moms' genome...
... but there's biology above and beyond the genes we get from mom (and not from dad). And that song is great for teaching us that the rest of the egg and the gestational experience in utero provide so much more to the development of the soon-to-be new human. So "slightly more than half of everything" is thanks to our mothers. Aw!
But, genetically, the mainstream idea is still that we're 50% our mom.
I teach very basic genetics because I teach evolution and anthropology.And I'm not (usually) a dummy.* I get it. It's a fact! I'm half, genetically, my mom and I'm also half my dad.
r = 0.5
Okay! But, given these facts about relatedness and how it's imagined in evolutionary biology, facts that I never ever questioned, I hope you can see why this report from 23andMe (personal genomics enterprise) blew my mind:
 |
| Percent similarity to Holly Dunsworth over 536070 SNPs (single nucleotide polymorphisms or, effectively/rather, a subset of known variants in the genome; Click on the image to enlarge). I am 85% like my mom and I am at least 76% like my students and friends who are sharing with me on 23andMe. Names of comparisons have been redacted. As far as I know, this kind of report is no longer offered by 23andMe. I spat back in 2011/12 and the platform has evolved since. |
Okay, first of all, it is a huge relief that, of all the people I'm sharing with on 23andMe, the one who squeezed me out of her body is the most genetically similar to me. Science works.
But that number there, with my mother, it is not 50%. It's quite a bit bigger than that. It says I'm over 85% the same as her.
What's more, I am also very similar to every single person I'm sharing with on the site, including example accounts from halfway around the world. Everyone is at least 60-ish% genetically similar to me, according to 23andMe. I know we're all "cousins," but my actual cousins are supposed to be 1/8th according to evolutionary biology. How can my mom be related to me by only one half? How can my actual cousins be only an eighth (which is 12.5%)?
What is up with evolutionary biology and this whole "r" thing?
Hi. Here is where, if they weren't already, people just got really annoyed with me. Evolutionary biology's "relatedness" or "r" is not the same as genetic similarity like that reported by 23andMe.
Okay!
But why not?
Let me help unpack the 85% genetic similarity with my mom. Remember, it's not because I'm inbred (which you have to take my word for, but notice that most everyone on there is over 70% genetically similar to me so...).
It's because my mom and dad, just like any two humans, share a lot in common genetically. Some of the alleles that I inherited from my dad are alleles that my mom inherited from her parents. So, not only is everything I got from her (50%) similar to her, but so are many of the parts that I got from my dad.
Let me get out my kid's arts supplies.
Here is a pretty common view of relatedness, genetically. In our imagination, parents are not related (r = 0) which can lead our imagination to think that their alleles are distinct. Here there are four distinct alleles/variants that could be passed onto offspring, with each offspring only getting one from mom and one from dad. In this case, the sperm carrying the orange variant and the egg with the blue variant made the baby.
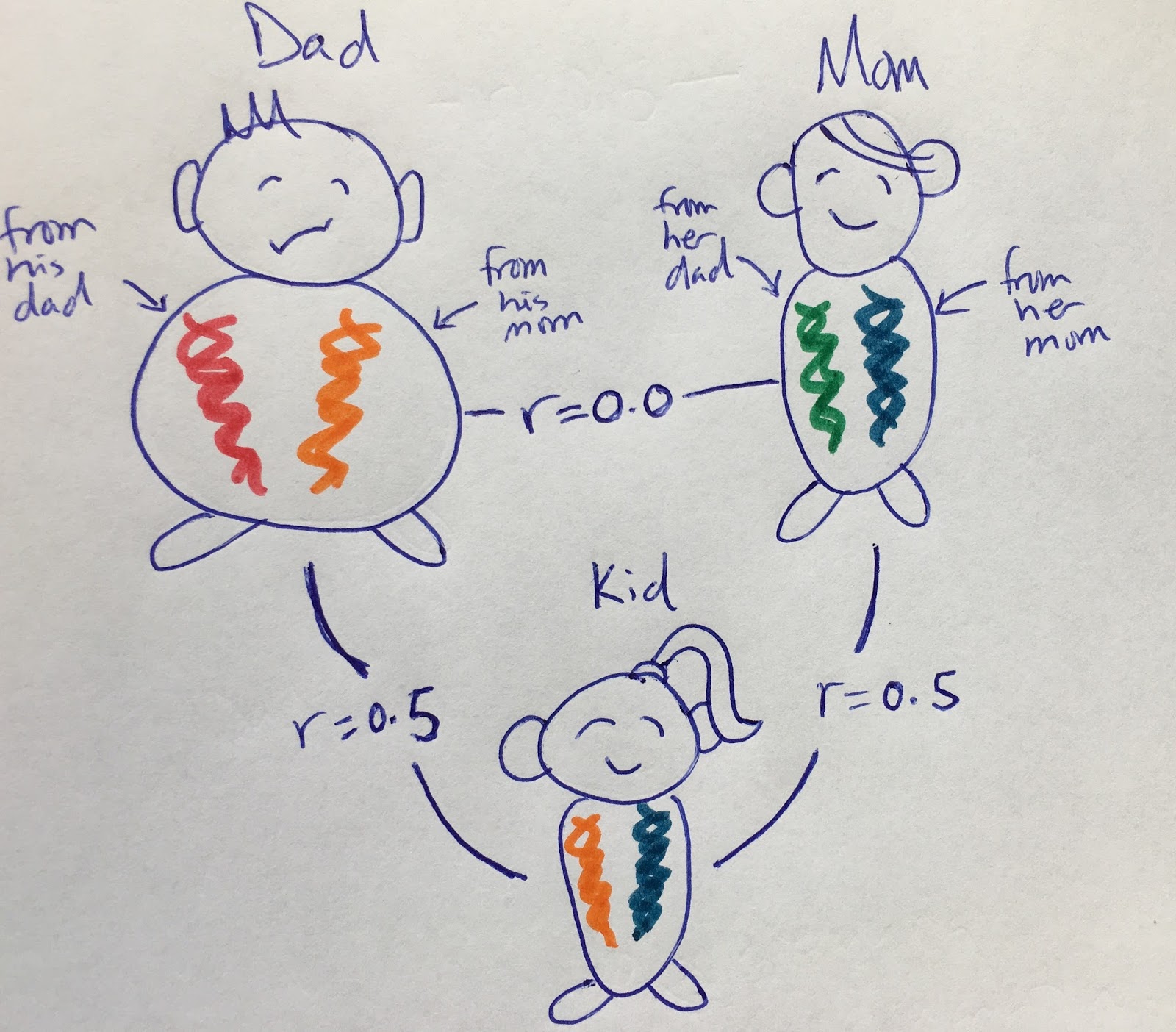 |
| 1. (Please, if you're horrified by the "r" business in these figures, read the post for explanation.) |
The green allele doesn't exist in the next example. As a result of there being only three variants for this gene or locus, mom and dad must share at least one allele, minimum. That means, they look related and that means that, depending on which egg and sperm make the kid, the kid could be more related to mom than to dad.
 |
| 2. (Please, if you're horrified by the "r" business in these figures, read the post for explanation.) |
Well, that's just insane. What does it matter whether she got the allele from mom or dad? I thought genes were selfish? (Sorry, for the outburst.)
Again, I realize I'm annoying people and probably much worse--like stomping all over theory and knowledge and science--by mixing up the different concepts of genetic similarity (e.g. 50%) with "r" (e.g. 0.5) and horribly misunderstanding all the nuance (and debate) about "r," but I'm doing it because I'm desperately trying to know why these two related ideas are, in fact, distinct.
One last pathetic cartoon.
In this third example, as is common in the genome, there are only two alleles/variants in existence (at an appreciable frequency, so not accounting for constant accumulation of de novo variation). An example of such a gene with only two known alleles is the "earwax gene" ABCC11 (there's a wet/waxy allele and dry/crumbly one). Here, the two alleles are orange and blue. Most humans in the species will have at least one allele in common with their mate for a gene with two alleles, and it's not because most humans are inbred, unless we want to redefine inbreeding to include very distant relatives (aside: which may be how the term is used by experts).
 |
| 3. (Please, if you're horrified by the "r" business in these figures, read the post for explanation.) |
And of course, making a kid with your same genotype is the only possible outcome if you and your mate are both homozygous (i.e. where both copies are of the same variant so that leaves no chance for variation in offspring unless there is a new mutation).
So, I wandered a little bit away from my point with these drawings, but I had to because I wanted to get down from where my imagination has me (us?) with "r" versus how things really are with reproduction. We are baby-making with vastly similar genomes to ours, so we are making babies with vastly similar genomes to ours.
So, I do see why biology says I'm related to my mom by one half. But, on the other hand, what does it matter if I got the thing I have in common with my mom from my mom or whether I got it from my dad? Because I got it. Period. It lives. Period.
[Inserted graf January 20, '17] Saying it matters where I got the similarity to my mom keeps us at r = 0.5. Saying it matters only that I inherited DNA like hers keeps us always, all of us, at r > 0.5 with our parents and our kids because any two babymakers share much of their genome.
And the fact that this (see 2 and 3) happens so often is why I'm a lot more than 50% genetically like my mom, and the same can be said about my genetic similarity to my dad without him even spitting for 23andMe.
So, here we are. I don't understand why our relatedness to one another, based on genetic similarity, is not "r."
I hope it's for really beautifully logical reasons and not something political.
Because...
If "r" was defined by genetic similarity, then would cooperating with my 76% genetically similar students and friends be more adaptive than the credit I currently get from evolutionary biology for cooperating with my own flesh and blood son?
If "r" was defined by genetic similarity, then could we use the power of math and theoretical biology to encourage broader cooperation among humans beyond their close kin?
So many questions.
Maybe I should re-learn the math and learn all the other math.
Nah. Not myself. At least, it wouldn't come fast enough for my appetite. Maybe someone who already knows the math could leave a comment and we could go from there...
And it would be worth it, you know, because despite my relatively weaker math skills, I bet we're more than 50% genetically similar.



